What is genetic material transferred to an organism?
![]()
Tree of life showing vertical and horizontal cistron transfers
Horizontal gene transfer (HGT) or lateral gene transfer (LGT)[1] [two] [3] is the motion of genetic fabric between unicellular and/or multicellular organisms other than by the ("vertical") manual of Dna from parent to offspring (reproduction).[4] HGT is an important factor in the evolution of many organisms.[five] [6]
Horizontal gene transfer is the primary mechanism for the spread of antibody resistance in bacteria,[v] [7] [8] [9] [10] and plays an of import office in the evolution of leaner that tin can degrade novel compounds such equally human-created pesticides[eleven] and in the evolution, maintenance, and manual of virulence.[12] It frequently involves temperate bacteriophages and plasmids.[xiii] [14] [15] Genes responsible for antibiotic resistance in ane species of bacteria tin can be transferred to another species of bacteria through various mechanisms of HGT such every bit transformation, transduction and conjugation, after arming the antibiotic resistant genes' recipient confronting antibiotics. The rapid spread of antibiotic resistance genes in this manner is becoming medically challenging to deal with. Ecological factors may likewise play a part in the HGT of antibiotic resistant genes.[xvi] It is also postulated that HGT promotes the maintenance of a universal life biochemistry and, later on, the universality of the genetic code.[17]
Most thinking in genetics has focused upon vertical transfer, but the importance of horizontal gene transfer among single-jail cell organisms is commencement to exist best-selling.[18] [19]
Factor delivery can exist seen as an artificial horizontal cistron transfer, and is a form of genetic engineering science.
History [edit]
Griffith'southward experiment, reported in 1928 past Frederick Griffith,[xx] was the first experiment suggesting that leaner are capable of transferring genetic information through a process known every bit transformation.[21] [22] Griffith'south findings were followed by research in the late 1930s and early 40s that isolated Deoxyribonucleic acid every bit the cloth that communicated this genetic information.
Horizontal genetic transfer was then described in Seattle in 1951, in a paper demonstrating that the transfer of a viral cistron into Corynebacterium diphtheriae created a virulent strain from a non-virulent strain,[23] also simultaneously solving the riddle of diphtheria (that patients could be infected with the bacteria but not have whatsoever symptoms, and and then suddenly catechumen later or never),[24] and giving the first example for the relevance of the lysogenic cycle.[25] Inter-bacterial cistron transfer was starting time described in Japan in a 1959 publication that demonstrated the transfer of antibody resistance betwixt different species of bacteria.[26] [27] In the mid-1980s, Syvanen[28] predicted that lateral gene transfer existed, had biological significance, and was involved in shaping evolutionary history from the start of life on World.
As Jian, Rivera and Lake (1999) put it: "Increasingly, studies of genes and genomes are indicating that considerable horizontal transfer has occurred betwixt prokaryotes"[29] (encounter also Lake and Rivera, 2007).[30] The phenomenon appears to have had some significance for unicellular eukaryotes as well. As Bapteste et al. (2005) observe, "additional evidence suggests that factor transfer might besides be an important evolutionary machinery in protist evolution."[31]
Grafting of 1 plant to some other tin can transfer chloroplasts (organelles in found cells that conduct photosynthesis), mitochondrial DNA, and the unabridged jail cell nucleus containing the genome to potentially make a new species.[32] Some Lepidoptera (e.g. monarch butterflies and silkworms) have been genetically modified by horizontal gene transfer from the wasp bracovirus.[33] Bites from insects in the family Reduviidae (assassin bugs) tin can, via a parasite, infect humans with the trypanosomal Chagas affliction, which can insert its Dna into the human genome.[34] It has been suggested that lateral factor transfer to humans from bacteria may play a role in cancer.[35]
Aaron Richardson and Jeffrey D. Palmer land: "Horizontal cistron transfer (HGT) has played a major role in bacterial evolution and is fairly common in certain unicellular eukaryotes. Notwithstanding, the prevalence and importance of HGT in the evolution of multicellular eukaryotes remain unclear."[36]
Due to the increasing amount of evidence suggesting the importance of these phenomena for evolution (encounter beneath) molecular biologists such as Peter Gogarten take described horizontal gene transfer every bit "A New Paradigm for Biology".[37]
Mechanisms [edit]
There are several mechanisms for horizontal gene transfer:[5] [38] [39]
- Transformation, the genetic amending of a cell resulting from the introduction, uptake and expression of foreign genetic material (DNA or RNA).[40] This process is relatively common in bacteria, but less so in eukaryotes.[41] Transformation is often used in laboratories to insert novel genes into bacteria for experiments or for industrial or medical applications. Come across also molecular biology and biotechnology.
- Transduction, the process in which bacterial Dna is moved from ane bacterium to some other by a virus (a bacteriophage, or phage).[40]
- Bacterial conjugation, a process that involves the transfer of DNA via a plasmid from a donor cell to a recombinant recipient cell during cell-to-cell contact.[40]
- Factor transfer agents, virus-like elements encoded past the host that are found in the alphaproteobacteria social club Rhodobacterales.[42]
Horizontal transposon transfer [edit]
A transposable element (TE) (also called a transposon or jumping gene) is a mobile segment of Dna that tin can sometimes pick upwardly a resistance factor and insert it into a plasmid or chromosome, thereby inducing horizontal factor transfer of antibiotic resistance.[40]
Horizontal transposon transfer (HTT) refers to the passage of pieces of Dna that are characterized past their ability to move from 1 locus to another between genomes by ways other than parent-to-offspring inheritance. Horizontal gene transfer has long been idea to exist crucial to prokaryotic evolution, merely in that location is a growing amount of data showing that HTT is a common and widespread phenomenon in eukaryote development as well.[43] On the transposable element side, spreading betwixt genomes via horizontal transfer may be viewed as a strategy to escape purging due to purifying selection, mutational disuse and/or host defense mechanisms.[44]
HTT tin occur with any type of transposable elements, just DNA transposons and LTR retroelements are more likely to exist capable of HTT because both have a stable, double-stranded DNA intermediate that is thought to be sturdier than the single-stranded RNA intermediate of non-LTR retroelements, which can exist highly degradable.[43] Non-autonomous elements may be less likely to transfer horizontally compared to autonomous elements considering they practice not encode the proteins required for their own mobilization. The structure of these non-autonomous elements generally consists of an intronless gene encoding a transposase poly peptide, and may or may non have a promoter sequence. Those that do not accept promoter sequences encoded within the mobile region rely on next host promoters for expression.[43] Horizontal transfer is idea to play an of import office in the TE life bike.[43]
HTT has been shown to occur betwixt species and across continents in both plants[45] and animals (Ivancevic et al. 2013), though some TEs have been shown to more than successfully colonize the genomes of certain species over others.[46] Both spatial and taxonomic proximity of species has been proposed to favor HTTs in plants and animals.[45] Information technology is unknown how the density of a population may affect the rate of HTT events within a population, but close proximity due to parasitism and cross contamination due to crowding accept been proposed to favor HTT in both plants and animals.[45] Successful transfer of a transposable chemical element requires delivery of DNA from donor to host prison cell (and to the germ line for multi-cellular organisms), followed by integration into the recipient host genome.[43] Though the actual mechanism for the transportation of TEs from donor cells to host cells is unknown, it is established that naked DNA and RNA can broadcast in bodily fluid.[43] Many proposed vectors include arthropods, viruses, freshwater snails (Ivancevic et al. 2013), endosymbiotic bacteria,[44] and intracellular parasitic bacteria.[43] In some cases, fifty-fifty TEs facilitate transport for other TEs.[46]
The arrival of a new TE in a host genome can have detrimental consequences because TE mobility may induce mutation. However, HTT tin also be beneficial past introducing new genetic material into a genome and promoting the shuffling of genes and TE domains among hosts, which can be co-opted past the host genome to perform new functions.[46] Moreover, transposition activity increases the TE re-create number and generates chromosomal rearrangement hotspots.[47] HTT detection is a difficult task because it is an ongoing miracle that is constantly changing in frequency of occurrence and composition of TEs inside host genomes. Furthermore, few species have been analyzed for HTT, making it difficult to plant patterns of HTT events between species. These problems tin pb to the underestimation or overestimation of HTT events between ancestral and current eukaryotic species.[47]
Methods of detection [edit]

A speciation event produces orthologs of a gene in the ii daughter species. A horizontal gene transfer event from one species to some other adds a xenolog of the gene to the receiving genome.
Horizontal gene transfer is typically inferred using bioinformatics methods, either past identifying atypical sequence signatures ("parametric" methods) or by identifying strong discrepancies between the evolutionary history of particular sequences compared to that of their hosts. The transferred gene (xenolog) plant in the receiving species is more closely related to the genes of the donor species than would exist expected.
Viruses [edit]
The virus called Mimivirus infects amoebae. Another virus, called Sputnik, also infects amoebae, but information technology cannot reproduce unless mimivirus has already infected the aforementioned cell.[48] "Sputnik's genome reveals further insight into its biological science. Although 13 of its genes show piffling similarity to whatsoever other known genes, three are closely related to mimivirus and mamavirus genes, perhaps cannibalized by the tiny virus every bit it packaged upward particles quondam in its history. This suggests that the satellite virus could perform horizontal cistron transfer between viruses, paralleling the way that bacteriophages ferry genes between bacteria."[49] Horizontal transfer is likewise seen betwixt geminiviruses and tobacco plants.[l]
Prokaryotes [edit]
Horizontal gene transfer is mutual among bacteria, even among very distantly related ones. This process is thought to exist a meaning cause of increased drug resistance[five] [51] when ane bacterial cell acquires resistance, and the resistance genes are transferred to other species.[52] [53] Transposition and horizontal cistron transfer, along with strong natural selective forces accept led to multi-drug resistant strains of Southward. aureus and many other pathogenic leaner.[forty] Horizontal factor transfer also plays a role in the spread of virulence factors, such as exotoxins and exoenzymes, amongst bacteria.[5] A prime example concerning the spread of exotoxins is the adaptive evolution of Shiga toxins in East. coli through horizontal gene transfer via transduction with Shigella species of bacteria.[54] Strategies to gainsay sure bacterial infections by targeting these specific virulence factors and mobile genetic elements take been proposed.[12] For example, horizontally transferred genetic elements play important roles in the virulence of East. coli, Salmonella, Streptococcus and Clostridium perfringens.[v]
In prokaryotes, restriction-modification systems are known to provide immunity against horizontal cistron transfer and in stabilizing mobile genetic elements. Genes encoding restriction modification systems have been reported to move between prokaryotic genomes within mobile genetic elements (MGE) such equally plasmids, prophages, insertion sequences/transposons, integrative conjugative elements (Water ice),[55] and integrons. Still, they are more ofttimes a chromosomal-encoded barrier to MGE than an MGE-encoded tool for cell infection.[56]
Lateral cistron transfer via a mobile genetic element, namely the integrated conjugative element (Water ice) Bs1 has been reported for its role in the global Deoxyribonucleic acid damage SOS response of the gram positive Bacillus subtilis.[57] Furthermore, it has been linked with the radiations and desiccation resistance of Bacillus pumilus SAFR-032 spores,[58] isolated from spacecraft cleanroom facilities.[59] [lx] [61]
Transposon insertion elements have been reported to increase the fettle of gram-negative E. coli strains through either major transpositions or genome rearrangements, and increasing mutation rates.[62] [63] In a study on the effects of long-term exposure of faux microgravity on not-pathogenic Eastward. coli, the results showed transposon insertions occur at loci, linked to SOS stress response.[64] When the same E. coli strain was exposed to a combination of simulated microgravity and trace (groundwork) levels of (the broad spectrum) antibiotic (chloramphenicol), the results showed transposon-mediated rearrangements (TMRs), disrupting genes involved in bacterial adhesion, and deleting an entire segment of several genes involved with motion and chemotaxis.[65] Both these studies take implications for microbial growth, adaptation to and antibiotic resistance in existent time space conditions.
Bacterial transformation [edit]
![]()
1: Donor leaner ii: Bacteria who volition receive the gene 3: The red portion represents the gene that will be transferred. Transformation in leaner happens in a certain environment.
Natural transformation is a bacterial adaptation for Deoxyribonucleic acid transfer (HGT) that depends on the expression of numerous bacterial genes whose products are responsible for this process.[66] [67] In general, transformation is a circuitous, energy-requiring developmental process. In social club for a bacterium to bind, take up and recombine exogenous DNA into its chromosome, it must become competent, that is, enter a special physiological state. Competence development in Bacillus subtilis requires expression of well-nigh forty genes.[68] The DNA integrated into the host chromosome is commonly (but with infrequent exceptions) derived from another bacterium of the aforementioned species, and is thus homologous to the resident chromosome. The chapters for natural transformation occurs in at least 67 prokaryotic species.[67] Competence for transformation is typically induced by loftier prison cell density and/or nutritional limitation, conditions associated with the stationary phase of bacterial growth. Competence appears to exist an adaptation for Deoxyribonucleic acid repair.[69] Transformation in bacteria can be viewed every bit a archaic sexual process, since it involves interaction of homologous DNA from two individuals to class recombinant Dna that is passed on to succeeding generations. Although transduction is the class of HGT nigh normally associated with bacteriophages, certain phages may also be able to promote transformation.[70]
Bacterial conjugation [edit]
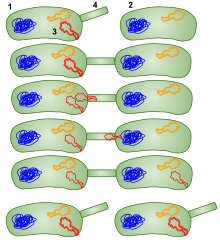
1: Donor leaner jail cell (F+ jail cell) 2: Leaner that receives the plasmid (F- prison cell) 3: Plasmid that will be moved to the other bacteria four: Pilus. Conjugation in bacteria using a sex pilus; and then the bacteria that received the plasmid can get give it to other bacteria besides.
Conjugation in Mycobacterium smegmatis, like conjugation in Eastward. coli, requires stable and extended contact between a donor and a recipient strain, is DNase resistant, and the transferred Dna is incorporated into the recipient chromosome by homologous recombination. However, unlike E. coli loftier frequency of recombination conjugation (Hfr), mycobacterial conjugation is a type of HGT that is chromosome rather than plasmid based.[71] Furthermore, in dissimilarity to East. coli (Hfr) conjugation, in M. smegmatis all regions of the chromosome are transferred with comparable efficiencies. Substantial blending of the parental genomes was plant as a event of conjugation, and this blending was regarded as reminiscent of that seen in the meiotic products of sexual reproduction.[71] [72]
Archaeal DNA transfer [edit]
The archaeon Sulfolobus solfataricus, when UV irradiated, strongly induces the formation of blazon IV pili which then facilitates cellular assemblage.[73] [74] Exposure to chemical agents that cause DNA impairment also induces cellular aggregation.[73] Other physical stressors, such as temperature shift or pH, do not induce aggregation, suggesting that Dna damage is a specific inducer of cellular assemblage.
UV-induced cellular aggregation mediates intercellular chromosomal HGT marker exchange with high frequency,[75] and UV-induced cultures display recombination rates that exceed those of uninduced cultures past every bit much as iii orders of magnitude. S. solfataricus cells aggregate preferentially with other cells of their own species.[75] Frols et al.[73] [76] and Ajon et al.[75] suggested that UV-inducible Dna transfer is likely an important machinery for providing increased repair of damaged DNA via homologous recombination. This process can exist regarded as a uncomplicated form of sexual interaction.
Another thermophilic species, Sulfolobus acidocaldarius, is able to undergo HGT. South. acidocaldarius tin can exchange and recombine chromosomal markers at temperatures up to 84 °C.[77] UV exposure induces pili formation and cellular aggregation.[75] Cells with the power to amass have greater survival than mutants lacking pili that are unable to aggregate. The frequency of recombination is increased past Deoxyribonucleic acid impairment induced by UV-irradiation[78] and by DNA damaging chemicals.[79]
The ups operon, containing five genes, is highly induced past UV irradiation. The proteins encoded past the ups operon are employed in UV-induced pili associates and cellular aggregation leading to intercellular DNA substitution and homologous recombination.[80] Since this system increases the fitness of S. acidocaldarius cells later on UV exposure, Wolferen et al.[80] [81] considered that transfer of Deoxyribonucleic acid likely takes place in order to repair UV-induced Dna damages by homologous recombination.
Eukaryotes [edit]
"Sequence comparisons propose recent horizontal transfer of many genes among diverse species including across the boundaries of phylogenetic 'domains'. Thus determining the phylogenetic history of a species can not exist done conclusively past determining evolutionary copse for single genes."[82]
Organelle to nuclear genome [edit]
- Assay of Deoxyribonucleic acid sequences suggests that horizontal gene transfer has occurred within eukaryotes from the chloroplast and mitochondrial genomes to the nuclear genome. As stated in the endosymbiotic theory, chloroplasts and mitochondria probably originated every bit bacterial endosymbionts of a progenitor to the eukaryotic cell.[83]
Organelle to organelle [edit]
- Mitochondrial genes moved to parasites of the Rafflesiaceae establish family from their hosts[84] [85] and from chloroplasts of a notwithstanding-unidentified plant to the mitochondria of the edible bean Phaseolus.[86]
Viruses to plants [edit]
- Plants are capable of receiving genetic information from viruses past horizontal gene transfer.[50]
Bacteria to fungi [edit]
- Horizontal transfer occurs from bacteria to some fungi, such equally the yeast Saccharomyces cerevisiae.[87]
Bacteria to plants [edit]
- Agrobacterium, a pathogenic bacterium that causes cells to proliferate as crown galls and proliferating roots is an example of a bacterium that can transfer genes to plants and this plays an important function in found evolution.[88]
Leaner to insects [edit]
- HhMAN1 is a gene in the genome of the coffee drupe borer (Hypothenemus hampei) that resembles bacterial genes, and is idea to be transferred from bacteria in the protrude'due south gut.[89] [90]
- oskar is an essential cistron for the specification of the germline in Holometabola and its origin is through to be due to a HGT issue followed by a fusion with a LOTUS domain.[91]
Bacteria to animals [edit]
- Bdelloid rotifers currently hold the 'record' for HGT in animals with ~viii% of their genes from bacterial origins.[92] Tardigrades were thought to intermission the tape with 17.5% HGT, merely that finding was an artifact of bacterial contamination.[93]
- A study establish the genomes of forty animals (including ten primates, four Caenorhabditis worms, and 12 Drosophila insects) independent genes which the researchers concluded had been transferred from bacteria and fungi by horizontal cistron transfer.[94] The researchers estimated that for some nematodes and Drosophila insects these genes had been acquired relatively recently.[95]
- A bacteriophage-mediated machinery transfers genes between prokaryotes and eukaryotes. Nuclear localization signals in bacteriophage terminal proteins (TP) prime Deoxyribonucleic acid replication and become covalently linked to the viral genome. The role of virus and bacteriophages in HGT in bacteria, suggests that TP-containing genomes could be a vehicle of inter-kingdom genetic information transference all throughout evolution.[96]
Endosymbiont to insects and nematodes [edit]
- The adzuki bean beetle has acquired genetic material from its (non-beneficial) endosymbiont Wolbachia.[97] New examples take recently been reported demonstrating that Wolbachia bacteria represent an important potential source of genetic material in arthropods and filarial nematodes.[98]
Found to found [edit]
- Striga hermonthica, a parasitic eudicot, has received a cistron from sorghum (Sorghum bicolor) to its nuclear genome.[99] The factor'southward functionality is unknown.
- A gene that immune ferns to survive in dark forests came from the hornwort, which grows in mats on streambanks or trees. The neochrome gene arrived about 180 meg years agone.[100]
Plants to animals [edit]
- The eastern emerald sea slug Elysia chlorotica has been suggested by fluorescence in situ hybridization (FISH) analysis to contain photosynthesis-supporting genes obtained from an algae (Vaucheria litorea) in their nutrition.[101] LGT in Sacoglossa is at present thought to exist an antiquity[102] and no trace of LGT was found upon sequencing the genome of Elysia chlorotica.[103]
- The whitefly Bemisia tabaci acquired a plant detoxification gene that neutralizes plant toxins.[104]
Establish to fungus [edit]
- Gene transfer between plants and fungi has been posited for a number of cases, including rice (Oryza sativa).
Fungi to insects [edit]
- Pea aphids (Acyrthosiphon pisum) contain multiple genes from fungi.[105] [106] Plants, fungi, and microorganisms tin can synthesize carotenoids, only torulene made past pea aphids is the just carotenoid known to be synthesized by an organism in the animal kingdom.[105]
Animals to animals [edit]
- Smelt fish received antifreeze poly peptide (AFP) gene from herring through a direct horizontal transfer.[107]
Man to protozoan [edit]
- The malaria pathogen Plasmodium vivax acquired genetic material from humans that might assist facilitate its long stay in the body.[108]
Human genome [edit]
- Ane study identified approximately 100 of humans' approximately 20,000 full genes which likely resulted from horizontal cistron transfer,[109] merely this number has been challenged past several researchers arguing these candidate genes for HGT are more probable the result of factor loss combined with differences in the rate of development.[110]
Artificial horizontal gene transfer [edit]
![]()
Earlier it is transformed, a bacterium is susceptible to antibiotics. A plasmid tin can be inserted when the leaner is under stress, and be incorporated into the bacterial DNA creating antibiotic resistance. When the plasmids are prepared they are inserted into the bacterial cell past either making pores in the plasma membrane with temperature extremes and chemical treatments, or making it semi permeable through the process of electrophoresis, in which electric currents create the holes in the membrane. Afterwards conditions return to normal the holes in the membrane close and the plasmids are trapped inside the bacteria where they get part of the genetic material and their genes are expressed by the bacteria.
Genetic engineering is essentially horizontal gene transfer, admitting with synthetic expression cassettes. The Sleeping Beauty transposon system[111] (SB) was developed as a synthetic gene transfer amanuensis that was based on the known abilities of Tc1/mariner transposons to invade genomes of extremely diverse species.[112] The SB organisation has been used to introduce genetic sequences into a wide variety of beast genomes.[113] [114]
Importance in evolution [edit]
Horizontal gene transfer is a potential confounding factor in inferring phylogenetic trees based on the sequence of one gene.[115] For example, given two distantly related bacteria that take exchanged a factor a phylogenetic tree including those species volition bear witness them to be closely related because that gene is the aforementioned even though well-nigh other genes are dissimilar. For this reason, it is oftentimes platonic to use other information to infer robust phylogenies such as the presence or absenteeism of genes or, more ordinarily, to include as wide a range of genes for phylogenetic analysis every bit possible.
For example, the about common gene to exist used for amalgam phylogenetic relationships in prokaryotes is the 16S ribosomal RNA gene since its sequences tend to exist conserved among members with close phylogenetic distances, but variable enough that differences can be measured. Still, in recent years it has too been argued that 16s rRNA genes can also be horizontally transferred. Although this may be infrequent, the validity of 16s rRNA-constructed phylogenetic trees must exist reevaluated.[116]
Biologist Johann Peter Gogarten suggests "the original metaphor of a tree no longer fits the information from recent genome research" therefore "biologists should utilise the metaphor of a mosaic to describe the dissimilar histories combined in private genomes and use the metaphor of a net to visualize the rich exchange and cooperative effects of HGT among microbes".[37] There exist several methods to infer such phylogenetic networks.
Using single genes every bit phylogenetic markers, it is difficult to trace organismal phylogeny in the presence of horizontal cistron transfer. Combining the simple coalescence model of cladogenesis with rare HGT horizontal gene transfer events advise there was no single most recent mutual ancestor that contained all of the genes ancestral to those shared among the iii domains of life. Each gimmicky molecule has its ain history and traces back to an private molecule cenancestor. However, these molecular ancestors were likely to exist present in unlike organisms at different times."[117]
Claiming to the tree of life [edit]
Horizontal gene transfer poses a possible claiming to the concept of the final universal mutual antecedent (LUCA) at the root of the tree of life first formulated past Carl Woese, which led him to propose the Archaea as a third domain of life.[118] Indeed, it was while examining the new iii-domain view of life that horizontal gene transfer arose equally a complicating upshot: Archaeoglobus fulgidus was seen as an anomaly with respect to a phylogenetic tree based upon the encoding for the enzyme HMGCoA reductase—the organism in question is a definite Archaean, with all the cell lipids and transcription machinery that are expected of an Archaean, but whose HMGCoA genes are of bacterial origin.[118] Scientists are broadly agreed on symbiogenesis, that mitochondria in eukaryotes derived from alpha-proteobacterial cells and that chloroplasts came from ingested blue-green alga, and other cistron transfers may take afflicted early eukaryotes. (In dissimilarity, multicellular eukaryotes have mechanisms to foreclose horizontal gene transfer, including separated germ cells.) If there had been continued and extensive cistron transfer, there would be a complex network with many ancestors, instead of a tree of life with sharply delineated lineages leading back to a LUCA.[118] [119] Nonetheless, a LUCA can exist identified, so horizontal transfers must have been relatively limited.[120]
Phylogenetic information in HGT [edit]
It has been remarked that, despite the complications, the detection of horizontal gene transfers brings valuable phylogenetic and dating information.[121]
The potential of HGT to be used for dating phylogenies has recently been confirmed.[122] [123]
The chromosomal organization of horizontal factor transfer [edit]
The acquisition of new genes has the potential to disorganize the other genetic elements and hinder the function of the bacterial jail cell, thus affecting the competitiveness of leaner. Consequently, bacterial adaptation lies in a conflict between the advantages of acquiring benign genes, and the need to maintain the organization of the balance of its genome. Horizontally transferred genes are typically concentrated in only ~ane% of the chromosome (in regions called hotspots). This concentration increases with genome size and with the charge per unit of transfer. Hotspots diversify by rapid gene turnover; their chromosomal distribution depends on local contexts (neighboring core genes), and content in mobile genetic elements. Hotspots concentrate most changes in cistron repertoires, reduce the trade-off between genome diversification and system, and should be treasure troves of strain-specific adaptive genes. Most mobile genetic elements and antibody resistance genes are in hotspots, simply many hotspots lack recognizable mobile genetic elements and exhibit frequent homologous recombination at flanking cadre genes. Overrepresentation of hotspots with fewer mobile genetic elements in naturally transformable leaner suggests that homologous recombination and horizontal cistron transfer are tightly linked in genome development.[124]
Genes [edit]
At that place is evidence for historical horizontal transfer of the following genes:
- Lycopene cyclase for carotenoid biosynthesis, between Chlorobi and Blue-green alga.[125]
- TetO gene conferring resistance to tetracycline, between Campylobacter jejuni.[126]
- Neochrome, a gene in some ferns that enhances their ability to survive in dim calorie-free. Believed to have been acquired from algae former during the Cretaceous.[127] [128]
- Transfer of a cysteine synthase from a bacterium into phytophagous mites and Lepidoptera assuasive the detoxification of cyanogenic glucosides produced by host plants.[129]
- The LINE1 sequence has transferred from humans to the gonorrhea bacteria.[130]
See likewise [edit]
- Agrobacterium, a bacterium well known for its ability to transfer DNA between itself and plants.
- Endogenous retrovirus
- Genetically modified organism
- Inferring horizontal gene transfer
- Integron
- Mobile genetic elements
- Phylogenetic network
- Phylogenetic tree
- Provirus
- Reassortment
- Retrotransposon
- Symbiogenesis
- Tree of life (biology)
- Xenobiology
References [edit]
- ^ Ochman H, Lawrence JG, Groisman EA (May 2000). "Lateral cistron transfer and the nature of bacterial innovation". Nature. 405 (6784): 299–304. Bibcode:2000Natur.405..299O. doi:10.1038/35012500. PMID 10830951. S2CID 85739173.
- ^ Dunning Hotopp JC (April 2011). "Horizontal gene transfer between bacteria and animals". Trends in Genetics. 27 (4): 157–63. doi:10.1016/j.tig.2011.01.005. PMC3068243. PMID 21334091.
- ^ Robinson KM, Sieber KB, Dunning Hotopp JC (Oct 2013). "A review of bacteria-animate being lateral factor transfer may inform our understanding of diseases like cancer". PLOS Genetics. ix (x): e1003877. doi:10.1371/periodical.pgen.1003877. PMC3798261. PMID 24146634.
- ^ Keeling PJ, Palmer JD (August 2008). "Horizontal gene transfer in eukaryotic evolution". Nature Reviews. Genetics. 9 (8): 605–eighteen. doi:x.1038/nrg2386. PMID 18591983. S2CID 213613.
- ^ a b c d due east f Gyles C, Boerlin P (March 2014). "Horizontally transferred genetic elements and their office in pathogenesis of bacterial affliction". Veterinary Pathology. 51 (2): 328–40. doi:10.1177/0300985813511131. PMID 24318976. S2CID 206510894.
- ^ Vaux F, Trewick SA, Morgan-Richards M (2017). "Speciation through the looking-glass". Biological Journal of the Linnean Society. 120 (2): 480–488. doi:10.1111/bij.12872.
- ^ OECD, Safety Assessment of Transgenic Organisms, Volume four: OECD Consensus Documents, 2010, pp.171-174
- ^ Kay E, Vogel TM, Bertolla F, Nalin R, Simonet P (July 2002). "In situ transfer of antibiotic resistance genes from transgenic (transplastomic) tobacco plants to leaner". Applied and Environmental Microbiology. 68 (7): 3345–51. Bibcode:2002ApEnM..68.3345K. doi:10.1128/aem.68.7.3345-3351.2002. PMC126776. PMID 12089013.
- ^ Koonin EV, Makarova KS, Aravind L (2001). "Horizontal gene transfer in prokaryotes: quantification and classification". Almanac Review of Microbiology. 55 (ane): 709–42. doi:10.1146/annurev.micro.55.1.709. PMC4781227. PMID 11544372.
- ^ Nielsen KM (1998). "Barriers to horizontal gene transfer by natural transformation in soil bacteria". APMIS. 84 (S84): 77–84. doi:10.1111/j.1600-0463.1998.tb05653.x. PMID 9850687. S2CID 26490197.
- ^ McGowan C, Fulthorpe R, Wright A, Tiedje JM (Oct 1998). "Evidence for interspecies factor transfer in the evolution of two,4-dichlorophenoxyacetic acid degraders". Applied and Ecology Microbiology. 64 (x): 4089–92. Bibcode:1998ApEnM..64.4089M. doi:10.1128/AEM.64.10.4089-4092.1998. PMC106609. PMID 9758850.
- ^ a b Great EC (December 2012). "Paradigms of pathogenesis: targeting the mobile genetic elements of affliction". Frontiers in Cellular and Infection Microbiology. ii: 161. doi:ten.3389/fcimb.2012.00161. PMC3522046. PMID 23248780.
- ^ Naik GA, Bhat LN, Chpoade BA, Lynch JM (1994). "Transfer of broad-host-range antibiotic resistance plasmids in soil microcosms". Curr. Microbiol. 28 (iv): 209–215. doi:ten.1007/BF01575963. S2CID 21015053.
- ^ Varga M, Kuntová L, Pantůček R, Mašlaňová I, Růžičková 5, Doškař J (July 2012). "Efficient transfer of antibody resistance plasmids by transduction within methicillin-resistant Staphylococcus aureus USA300 clone". FEMS Microbiology Messages. 332 (2): 146–52. doi:ten.1111/j.1574-6968.2012.02589.x. PMID 22553940.
- ^ Varga M, Pantu Ček R, Ru Žičková V, Doškař J (January 2016). "Molecular characterization of a new efficiently transducing bacteriophage identified in meticillin-resistant Staphylococcus aureus". The Journal of General Virology. 97 (one): 258–268. doi:10.1099/jgv.0.000329. PMID 26537974.
- ^ Cairns J, Ruokolainen L, Hultman J, Tamminen 1000, Virta G, Hiltunen T (2018-04-nineteen). "Ecology determines how depression antibody concentration impacts customs composition and horizontal transfer of resistance genes". Communications Biology. i (1): 35. doi:x.1038/s42003-018-0041-seven. PMC6123812. PMID 30271921.
- ^ Kubyshkin V, Acevedo-Rocha CG, Budisa Due north (February 2018). "On universal coding events in protein biogenesis". Bio Systems. 164: 16–25. doi:10.1016/j.biosystems.2017.x.004. PMID 29030023.
- ^ Lin Edwards (Oct 4, 2010). "Horizontal factor transfer in microbes much more frequent than previously thought". PhysOrg.com. Retrieved 2012-01-06 .
- ^ Arnold C (April 2011). "To share and share alike". Scientific American. 304 (4): thirty–1. Bibcode:2011SciAm.304d..30A. doi:10.1038/scientificamerican0411-30. PMID 21495477.
- ^ Griffith F (January 1928). "The Significance of Pneumococcal Types". The Journal of Hygiene. Cambridge University Printing. 27 (2): 113–59. doi:ten.1017/S0022172400031879. JSTOR 4626734. PMC2167760. PMID 20474956.
- ^ Lorenz MG, Wackernagel Westward (September 1994). "Bacterial factor transfer by natural genetic transformation in the surroundings". Microbiological Reviews. 58 (3): 563–602. doi:10.1128/MMBR.58.3.563-602.1994. PMC372978. PMID 7968924.
- ^ Downie AW (November 1972). "Pneumococcal transformation--a backward view. 4th Griffith Memorial Lecture" (PDF). Journal of General Microbiology. 73 (1): one–eleven. doi:ten.1099/00221287-73-1-1. PMID 4143929.
- ^ Freeman VJ (June 1951). "Studies on the virulence of bacteriophage-infected strains of Corynebacterium diphtheriae". Periodical of Bacteriology. 61 (6): 675–88. doi:ten.1128/JB.61.half-dozen.675-688.1951. PMC386063. PMID 14850426.
- ^ Phillip Marguilies "Epidemics: Deadly diseases throughout history". Rosen, New York. 2005.
- ^ André Lwoff (1965). "Interaction among Virus, Cell, and Organism". Nobel Lecture for the Nobel Prize in Physiology or Medicine.
- ^ Ochiai Thou, Yamanaka T, Kimura K, Sawada O (1959). "Inheritance of drug resistance (and its transfer) betwixt Shigella strains and Betwixt Shigella and E. coli strains". Hihon Iji Shimpor (in Japanese). 1861: 34.
- ^ Akiba T, Koyama K, Ishiki Y, Kimura S, Fukushima T (April 1960). "On the machinery of the development of multiple-drug-resistant clones of Shigella". Japanese Periodical of Microbiology. 4 (2): 219–27. doi:x.1111/j.1348-0421.1960.tb00170.x. PMID 13681921.
- ^ Syvanen M (January 1985). "Cross-species factor transfer; implications for a new theory of development" (PDF). Journal of Theoretical Biology. 112 (ii): 333–43. Bibcode:1985JThBi.112..333S. doi:10.1016/S0022-5193(85)80291-v. PMID 2984477.
- ^ Jain R, Rivera MC, Lake JA (March 1999). "Horizontal factor transfer among genomes: the complexity hypothesis". Proceedings of the National Academy of Sciences of the United states of America. 96 (vii): 3801–half dozen. Bibcode:1999PNAS...96.3801J. doi:10.1073/pnas.96.7.3801. PMC22375. PMID 10097118.
- ^ Rivera MC, Lake JA (September 2004). "The band of life provides evidence for a genome fusion origin of eukaryotes" (PDF). Nature. 431 (7005): 152–5. Bibcode:2004Natur.431..152R. doi:x.1038/nature02848. PMID 15356622. S2CID 4349149. Archived from the original (PDF) on 2007-09-27.
- ^ Bapteste E, Susko E, Leigh J, MacLeod D, Charlebois RL, Doolittle WF (May 2005). "Do orthologous gene phylogenies really back up tree-thinking?". BMC Evolutionary Biology. 5 (ane): 33. doi:10.1186/1471-2148-5-33. PMC1156881. PMID 15913459.
- ^ Le Folio M (2016-03-17). "Farmers may have been accidentally making GMOs for millennia". The New Scientist. Retrieved 2016-07-eleven .
- ^ Gasmi 50, Boulain H, Gauthier J, Hua-Van A, Musset M, Jakubowska AK, et al. (September 2015). "Recurrent Domestication past Lepidoptera of Genes from Their Parasites Mediated by Bracoviruses". PLOS Genetics. 11 (9): e1005470. doi:ten.1371/journal.pgen.1005470. PMC4574769. PMID 26379286.
- ^ Yong Eastward (2010-02-14). "Genes from Chagas parasite tin can transfer to humans and be passed on to children". National Geographic. Retrieved 2016-07-thirteen .
- ^ Riley DR, Sieber KB, Robinson KM, White JR, Ganesan A, Nourbakhsh S, Dunning Hotopp JC (2013). "Leaner-human somatic cell lateral gene transfer is enriched in cancer samples". PLOS Computational Biological science. ix (six): e1003107. Bibcode:2013PLSCB...9E3107R. doi:ten.1371/journal.pcbi.1003107. PMC3688693. PMID 23840181.
- ^ Richardson AO, Palmer JD (2007). "Horizontal gene transfer in plants" (PDF). Journal of Experimental Botany. 58 (1): 1–nine. doi:ten.1093/jxb/erl148. PMID 17030541. Archived from the original (PDF) on 2007-09-27.
- ^ a b Gogarten, Peter (2000). "Horizontal Gene Transfer: A New Paradigm for Biology". Esalen Eye for Theory and Research Conference . Retrieved 2007-03-eighteen .
- ^ Kenneth Todar. "Bacterial Resistance to Antibiotics". The Microbial World: Lectures in Microbiology, Section of Bacteriology, Academy of Wisconsin-Madison. Archived from the original on Jan 15, 2012. Retrieved January 6, 2012.
- ^ Stanley Maloy (July 15, 2002). "Horizontal Factor Transfer". San Diego State University. Retrieved January 6, 2012.
- ^ a b c d e Stearns, S. C., & Hoekstra, R. F. (2005). Evolution: An introduction (2nd ed.). Oxford, NY: Oxford Univ. Press. pp. 38-40.
- ^ Renner SS, Bellot S (2012). "Horizontal Cistron Transfer in Eukaryotes: Fungi-to-Plant and Plant-to-Institute Transfers of Organellar Deoxyribonucleic acid". Genomics of Chloroplasts and Mitochondria. Advances in Photosynthesis and Respiration. Vol. 35. Springer Scientific discipline+Business Media B.V. pp. 223–235. doi:10.1007/978-94-007-2920-9_10. ISBN978-94-007-2919-3.
- ^ Maxmen A (2010). "Virus-like particles speed bacterial evolution". Nature. doi:ten.1038/news.2010.507.
- ^ a b c d e f chiliad Schaack S, Gilbert C, Feschotte C (September 2010). "Promiscuous Dna: horizontal transfer of transposable elements and why it matters for eukaryotic evolution". Trends in Environmental & Evolution. 25 (9): 537–46. doi:10.1016/j.tree.2010.06.001. PMC2940939. PMID 20591532.
- ^ a b Dupeyron M, Leclercq S, Cerveau Due north, Bouchon D, Gilbert C (January 2014). "Horizontal transfer of transposons between and inside crustaceans and insects". Mobile Deoxyribonucleic acid. five (one): iv. doi:10.1186/1759-8753-five-4. PMC3922705. PMID 24472097.
- ^ a b c El Baidouri Grand, Carpentier MC, Cooke R, Gao D, Lasserre Eastward, Llauro C, et al. (May 2014). "Widespread and frequent horizontal transfers of transposable elements in plants". Genome Inquiry. 24 (v): 831–8. doi:10.1101/gr.164400.113. PMC4009612. PMID 24518071.
- ^ a b c Ivancevic AM, Walsh AM, Kortschak RD, Adelson DL (Dec 2013). "Jumping the fine LINE between species: horizontal transfer of transposable elements in animals catalyses genome evolution". BioEssays. 35 (12): 1071–82. doi:10.1002/bies.201300072. PMID 24003001. S2CID 6968210.
- ^ a b Wallau GL, Ortiz MF, Loreto EL (2012). "Horizontal transposon transfer in eukarya: detection, bias, and perspectives". Genome Biology and Development. 4 (viii): 689–99. doi:ten.1093/gbe/evs055. PMC3516303. PMID 22798449.
- ^ La Scola B, Desnues C, Pagnier I, Robert C, Barrassi Fifty, Fournous G, et al. (September 2008). "The virophage as a unique parasite of the giant mimivirus". Nature. 455 (7209): 100–iv. Bibcode:2008Natur.455..100L. doi:x.1038/nature07218. PMID 18690211. S2CID 4422249.
- ^ Pearson H (August 2008). "'Virophage' suggests viruses are alive". Nature. 454 (7205): 677. Bibcode:2008Natur.454..677P. doi:x.1038/454677a. PMID 18685665.
- ^ a b Bejarano ER, Khashoggi A, Witty Thou, Lichtenstein C (January 1996). "Integration of multiple repeats of geminiviral DNA into the nuclear genome of tobacco during evolution". Proceedings of the National Academy of Sciences of the Usa of America. 93 (2): 759–64. Bibcode:1996PNAS...93..759B. doi:ten.1073/pnas.93.ii.759. PMC40128. PMID 8570630.
- ^ Barlow M (2009). "What antimicrobial resistance has taught us nigh horizontal gene transfer". Horizontal Gene Transfer. Methods in Molecular Biology. Vol. 532. pp. 397–411. doi:10.1007/978-1-60327-853-9_23. ISBN978-1-60327-852-2. PMID 19271198.
- ^ Hawkey PM, Jones AM (September 2009). "The changing epidemiology of resistance". The Periodical of Antimicrobial Chemotherapy. 64 Suppl 1 (Suppl i): i3-ten. doi:10.1093/jac/dkp256. PMID 19675017.
- ^ Francino MP, ed. (2012). Horizontal Cistron Transfer in Microorganisms. Caister Academic Press. ISBN978-ane-908230-10-2.
- ^ Strauch E, Lurz R, Beutin 50 (December 2001). "Characterization of a Shiga toxin-encoding temperate bacteriophage of Shigella sonnei". Infection and Immunity. 69 (12): 7588–95. doi:ten.1128/IAI.69.12.7588-7595.2001. PMC98851. PMID 11705937.
- ^ Johnson CM, Grossman Ad (Nov 2015). "Integrative and Conjugative Elements (ICEs): What They Exercise and How They Work". Annual Review of Genetics. 42 (1): 577–601. doi:10.1146/annurev-genet-112414-055018. PMC5180612. PMID 26473380.
- ^ Oliveira PH, Touchon G, Rocha EP (September 2014). "The interplay of restriction-modification systems with mobile genetic elements and their prokaryotic hosts". Nucleic Acids Enquiry. 49 (16): 10618–10631. doi:10.1093/nar/gku734. PMC4176335. PMID 25120263.
- ^ Auchtung JM, Lee CA, Garrison KL, Grossman Ad (June 2007). "Identification and label of the immunity repressor (ImmR) that controls the mobile genetic element ICE Bs1 of Bacillus subtilis". PLOS Genet. 64 (6): 1515–1528. doi:x.1111/j.1365-2958.2007.05748.x. PMC3320793. PMID 17511812.
- ^ Tirumalai MR, Fox GE (September 2013). "An IceBs1-like element may be associated with the extreme radiation and desiccation resistance of Bacillus pumilus SAFR-032 spores". Extremophiles. 17 (five): 767–774. doi:x.1007/s00792-013-0559-z. PMID 23812891. S2CID 8675124.
- ^ Link L, Sawyer J, Venkateswaran K, Nicholson W (February 2004). "Extreme spore UV resistance of Bacillus pumilus isolates obtained from an ultraclean Spacecraft Assembly Facility". Microb Ecol. 47 (ii): 159–163. doi:10.1007/s00248-003-1029-4. PMID 14502417. S2CID 13416635.
- ^ Newcombe DA, Schuerger AC, Benardini JN, Dickinson D, Tanner R, Venkateswaran K (December 2005). "Survival of spacecraft-associated microorganisms under fake martian UV irradiation". Appl Environ Microbiol. 71 (12): 8147–8156. Bibcode:2005ApEnM..71.8147N. doi:x.1128/AEM.71.12.8147-8156.2005. PMC1317311. PMID 16332797.
- ^ Kempf MJ, Chen F, Kern R, Venkateswaran K (June 2005). "Recurrent isolation of hydrogen peroxide-resistant spores of Bacillus pumilus from a spacecraft assembly facility". Astrobiology. 5 (3): 391–405. Bibcode:2005AsBio...5..391K. doi:10.1089/ast.2005.5.391. PMID 15941382.
- ^ Biel SW, Hartl DL (June 1983). "Evolution of transposons: natural option for Tn5 in Escherichia coli K12". Genetics. 103 (4): 581–592. doi:10.1093/genetics/103.4.581. PMC1202041. PMID 6303898.
- ^ Chao L, Vargas C, Spear BB, Cox EC (1983). "Transposable elements as mutator genes in evolution". Nature. 303 (5918): 633–635. Bibcode:1983Natur.303..633C. doi:10.1038/303633a0. PMC1202041. PMID 6303898.
- ^ Tirumalai MR, Karouia F, Tran Q, Stepanov VG, Bruce RJ, Ott M, Pierson DL, Pull a fast one on GE (May 2017). "The accommodation of Escherichia coli cells grown in simulated microgravity for an extended flow is both phenotypic and genomic". NPJ Microgravity. 3 (fifteen): 15. doi:10.1038/s41526-017-0020-1. PMC5460176. PMID 28649637.
- ^ Tirumalai MR, Karouia F, Tran Q, Stepanov VG, Bruce RJ, Ott M, Pierson DL, Fox GE (January 2019). "Evaluation of acquired antibiotic resistance in Escherichia coli exposed to long-term low-shear modeled microgravity and background antibody exposure". mBio. 10 (e02637-18). doi:10.1128/mBio.02637-18. PMC6336426. PMID 30647159.
- ^ Chen I, Dubnau D (March 2004). "DNA uptake during bacterial transformation". Nature Reviews. Microbiology. ii (3): 241–9. doi:ten.1038/nrmicro844. PMID 15083159. S2CID 205499369.
- ^ a b Johnsborg O, Eldholm V, Håvarstein LS (December 2007). "Natural genetic transformation: prevalence, mechanisms and function". Research in Microbiology. 158 (ten): 767–78. doi:10.1016/j.resmic.2007.09.004. PMID 17997281.
- ^ Solomon JM, Grossman Advertising (Apr 1996). "Who'due south competent and when: regulation of natural genetic competence in bacteria". Trends in Genetics. 12 (4): 150–five. doi:10.1016/0168-9525(96)10014-vii. PMID 8901420.
- ^ Michod RE, Bernstein H, Nedelcu AM (May 2008). "Adaptive value of sex in microbial pathogens" (PDF). Infection, Genetics and Evolution. 8 (iii): 267–85. doi:10.1016/j.meegid.2008.01.002. PMID 18295550.
- ^ Great EC, Bliskovsky VV, Malagon F, Baker JD, Prince JS, Klaus JS, Adhya SL (January 2017). "Novel "Superspreader" Bacteriophages Promote Horizontal Gene Transfer by Transformation". mBio. 8 (1): e02115-16. doi:10.1128/mBio.02115-xvi. PMC5241400. PMID 28096488.
- ^ a b Gray TA, Krywy JA, Harold J, Palumbo MJ, Derbyshire KM (July 2013). "Distributive bridal transfer in mycobacteria generates progeny with meiotic-like genome-wide mosaicism, allowing mapping of a mating identity locus". PLOS Biological science. 11 (seven): e1001602. doi:10.1371/journal.pbio.1001602. PMC3706393. PMID 23874149.
- ^ Derbyshire KM, Gray TA (2014). "Distributive Bridal Transfer: New Insights into Horizontal Cistron Transfer and Genetic Commutation in Mycobacteria". Microbiology Spectrum. ii (one): 61–79. doi:x.1128/microbiolspec.MGM2-0022-2013. PMC4259119. PMID 25505644.
- ^ a b c Fröls S, Ajon K, Wagner 1000, Teichmann D, Zolghadr B, Folea M, et al. (November 2008). "UV-inducible cellular aggregation of the hyperthermophilic archaeon Sulfolobus solfataricus is mediated by pili formation" (PDF). Molecular Microbiology. seventy (4): 938–52. doi:10.1111/j.1365-2958.2008.06459.10. PMID 18990182.
- ^ Allers T (November 2011). "Swapping genes to survive - a new office for archaeal type IV pili". Molecular Microbiology. 82 (4): 789–91. doi:10.1111/j.1365-2958.2011.07860.x. PMID 21992544.
- ^ a b c d Ajon M, Fröls Southward, van Wolferen 1000, Stoecker K, Teichmann D, Driessen AJ, et al. (November 2011). "UV-inducible DNA substitution in hyperthermophilic archaea mediated past type Iv pili" (PDF). Molecular Microbiology. 82 (iv): 807–17. doi:ten.1111/j.1365-2958.2011.07861.10. PMID 21999488.
- ^ Fröls S, White MF, Schleper C (February 2009). "Reactions to UV damage in the model archaeon Sulfolobus solfataricus". Biochemical Society Transactions. 37 (Pt 1): 36–41. doi:x.1042/BST0370036. PMID 19143598.
- ^ Grogan DW (June 1996). "Exchange of genetic markers at extremely loftier temperatures in the archaeon Sulfolobus acidocaldarius". Journal of Bacteriology. 178 (11): 3207–xi. doi:10.1128/jb.178.11.3207-3211.1996. PMC178072. PMID 8655500.
- ^ Wood ER, Ghané F, Grogan DW (September 1997). "Genetic responses of the thermophilic archaeon Sulfolobus acidocaldarius to short-wavelength UV calorie-free". Journal of Bacteriology. 179 (18): 5693–8. doi:x.1128/jb.179.eighteen.5693-5698.1997. PMC179455. PMID 9294423.
- ^ Reilly MS, Grogan DW (Feb 2002). "Biological furnishings of DNA damage in the hyperthermophilic archaeon Sulfolobus acidocaldarius". FEMS Microbiology Messages. 208 (ane): 29–34. doi:x.1016/s0378-1097(01)00575-4. PMID 11934490.
- ^ a b van Wolferen G, Ajon M, Driessen AJ, Albers SV (Dec 2013). "Molecular analysis of the UV-inducible pili operon from Sulfolobus acidocaldarius". MicrobiologyOpen. 2 (vi): 928–37. doi:10.1002/mbo3.128. PMC3892339. PMID 24106028.
- ^ van Wolferen K, Ma X, Albers SV (September 2015). "DNA Processing Proteins Involved in the UV-Induced Stress Response of Sulfolobales". Journal of Bacteriology. 197 (18): 2941–51. doi:10.1128/JB.00344-15. PMC4542170. PMID 26148716.
- ^ Melcher U (2001). "Molecular genetics: Horizontal factor transfer". Stillwater, Oklahoma USA: Oklahoma Land University. Archived from the original on 2016-03-04. Retrieved 2015-08-20 .
- ^ Blanchard JL, Lynch M (July 2000). "Organellar genes: why exercise they stop up in the nucleus?". Trends in Genetics. 16 (seven): 315–xx. doi:ten.1016/S0168-9525(00)02053-nine. PMID 10858662. Discusses theories on how mitochondria and chloroplast genes are transferred into the nucleus, and also what steps a gene needs to go through in order to complete this process.
- ^ Davis CC, Wurdack KJ (July 2004). "Host-to-parasite gene transfer in flowering plants: phylogenetic testify from Malpighiales". Science. 305 (5684): 676–8. Bibcode:2004Sci...305..676D. doi:10.1126/science.1100671. PMID 15256617. S2CID 16180594.
- ^ Nickrent DL, Blarer A, Qiu YL, Vidal-Russell R, Anderson Atomic number 26 (Oct 2004). "Phylogenetic inference in Rafflesiales: the influence of rate heterogeneity and horizontal factor transfer". BMC Evolutionary Biology. 4 (1): 40. doi:10.1186/1471-2148-4-40. PMC528834. PMID 15496229.
- ^ Woloszynska M, Bocer T, Mackiewicz P, Janska H (November 2004). "A fragment of chloroplast Dna was transferred horizontally, probably from non-eudicots, to mitochondrial genome of Phaseolus". Found Molecular Biology. 56 (5): 811–xx. doi:10.1007/s11103-004-5183-y. PMID 15803417. S2CID 14198321.
- ^ Hall C, Brachat S, Dietrich FS (June 2005). "Contribution of horizontal gene transfer to the evolution of Saccharomyces cerevisiae". Eukaryotic Cell. iv (vi): 1102–15. doi:10.1128/EC.4.half-dozen.1102-1115.2005. PMC1151995. PMID 15947202.
- ^ Quispe-Huamanquispe DG, Gheysen G, Kreuze JF (2017). "Agrobacterium T-DNAs". Frontiers in Plant Science. eight: 2015. doi:ten.3389/fpls.2017.02015. PMC5705623. PMID 29225610.
- ^ Lee Phillips M (2012). "Bacterial cistron helps java protrude go its ready". Nature. doi:x.1038/nature.2012.10116. S2CID 211729274.
- ^ Acuña R, Padilla BE, Flórez-Ramos CP, Rubio JD, Herrera JC, Benavides P, et al. (March 2012). "Adaptive horizontal transfer of a bacterial cistron to an invasive insect pest of coffee". Proceedings of the National University of Sciences of the United states of america of America. 109 (xi): 4197–202. Bibcode:2012PNAS..109.4197A. doi:x.1073/pnas.1121190109. PMC3306691. PMID 22371593.
- ^ Blondel L, Jones ET, Extavour GC (February 2020). "Bacterial contribution to genesis of the novel germ line determinant oskar". eLife. 24 (nine): e45539. doi:x.7554/eLife.45539. PMC7250577. PMID 32091394.
- ^ Traci Watson (fifteen Nov 2012). "Bdelloids Surviving on Borrowed DNA". Science/AAAS News.
- ^ Koutsovoulos G, Kumar S, Laetsch DR, Stevens 50, Daub J, Conlon C, et al. (May 2016). "No testify for all-encompassing horizontal gene transfer in the genome of the tardigrade Hypsibius dujardini". Proceedings of the National Academy of Sciences of the Us of America. 113 (xviii): 5053–viii. Bibcode:2016PNAS..113.5053K. doi:10.1073/pnas.1600338113. PMC4983863. PMID 27035985.
- ^ Crisp A, Boschetti C, Perry Thou, Tunnacliffe A, Micklem G (March 2015). "Expression of multiple horizontally caused genes is a hallmark of both vertebrate and invertebrate genomes". Genome Biology. 16: 50. doi:10.1186/s13059-015-0607-3. PMC4358723. PMID 25785303.
- ^ Madhusoodanan J (2015-03-12). "Horizontal Cistron Transfer a Hallmark of Animal Genomes?". The Scientist . Retrieved 2016-07-xiv .
- ^ Redrejo-Rodríguez M, Muñoz-Espín D, Holguera I, Mencía M, Salas M (Nov 2012). "Functional eukaryotic nuclear localization signals are widespread in terminal proteins of bacteriophages". Proceedings of the National Academy of Sciences of the The states. 109 (45): 18482–7. Bibcode:2012PNAS..10918482R. doi:10.1073/pnas.1216635109. PMC3494942. PMID 23091024.
- ^ Kondo N, Nikoh N, Ijichi N, Shimada Yard, Fukatsu T (October 2002). "Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect". Proceedings of the National Academy of Sciences of the The states of America. 99 (22): 14280–5. Bibcode:2002PNAS...9914280K. doi:ten.1073/pnas.222228199. PMC137875. PMID 12386340.
- ^ Dunning Hotopp JC, Clark ME, Oliveira DC, Foster JM, Fischer P, Muñoz Torres MC, et al. (September 2007). "Widespread lateral gene transfer from intracellular bacteria to multicellular eukaryotes". Science. 317 (5845): 1753–6. Bibcode:2007Sci...317.1753H. doi:10.1126/scientific discipline.1142490. PMID 17761848. S2CID 10787254.
- ^ Yoshida Southward, Maruyama South, Nozaki H, Shirasu Thousand (May 2010). "Horizontal factor transfer by the parasitic plant Striga hermonthica". Science. 328 (5982): 1128. Bibcode:2010Sci...328.1128Y. doi:10.1126/science.1187145. PMID 20508124. S2CID 39376164.
- ^ Carl Zimmer (Apr 17, 2014). "Plants That Do Genetic Engineering". New York Times.
- ^ Schwartz JA, Curtis NE, Pierce SK (December 2014). "FISH labeling reveals a horizontally transferred algal (Vaucheria litorea) nuclear gene on a sea slug (Elysia chlorotica) chromosome". The Biological Message. 227 (3): 300–12. doi:x.1086/BBLv227n3p300. PMID 25572217. S2CID 21742354.
- ^ Rauch C, Vries J, Rommel Southward, Rose LE, Woehle C, Christa G, et al. (Baronial 2015). "Why It Is Time to Expect Beyond Algal Genes in Photosynthetic Slugs". Genome Biology and Evolution. 7 (9): 2602–7. doi:x.1093/gbe/evv173. PMC4607529. PMID 26319575.
- ^ Bhattacharya D, Pelletreau KN, Price DC, Sarver KE, Rumpho ME (August 2013). "Genome assay of Elysia chlorotica Egg Deoxyribonucleic acid provides no prove for horizontal gene transfer into the germ line of this Kleptoplastic Mollusc". Molecular Biological science and Evolution. xxx (8): 1843–52. doi:10.1093/molbev/mst084. PMC3708498. PMID 23645554.
- ^ https://world wide web.cell.com/cell/pdf/S0092-8674(21)00164-1.pdf
- ^ a b Moran NA, Jarvik T (Apr 2010). "Lateral transfer of genes from fungi underlies carotenoid product in aphids". Science. 328 (5978): 624–7. Bibcode:2010Sci...328..624M. doi:ten.1126/science.1187113. PMID 20431015. S2CID 14785276.
- ^ Fukatsu T (April 2010). "Evolution. A fungal past to insect colour". Science. 328 (5978): 574–5. Bibcode:2010Sci...328..574F. doi:x.1126/science.1190417. PMID 20431000. S2CID 23686682.
- ^ Wilcox, Christie (2021-06-09). "Deoxyribonucleic acid Jumps Between Animal Species. No One Knows How Oftentimes". Quanta Magazine . Retrieved 2021-06-fifteen .
- ^ Bar D (16 February 2011). "Evidence of Massive Horizontal Gene Transfer Between Humans and Plasmodium vivax". Nature Precedings. doi:x.1038/npre.2011.5690.i.
- ^ "Human beings' ancestors have routinely stolen genes from other species". The Economist. 14 March 2015. Retrieved 17 March 2015.
- ^ Salzberg SL, White O, Peterson J, Eisen JA (June 2001). "Microbial genes in the human being genome: lateral transfer or gene loss?". Science. 292 (5523): 1903–six. Bibcode:2001Sci...292.1903S. doi:10.1126/science.1061036. PMID 11358996. S2CID 17016011.
- ^ Ivics Z, Hackett PB, Plasterk RH, Izsvák Z (November 1997). "Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in man cells". Cell. 91 (four): 501–10. doi:10.1016/S0092-8674(00)80436-5. PMID 9390559. S2CID 17908472.
- ^ Plasterk RH (1996). "The Tc1/mariner transposon family". In Saedler H, Gierl A (eds.). Transposable Elements. Electric current Topics in Microbiology and Immunology. Vol. 204. pp. 125–143. doi:x.1007/978-3-642-79795-8_6. ISBN978-3-642-79797-two. PMID 8556864.
- ^ Izsvák Z, Ivics Z, Plasterk RH (September 2000). "Sleeping Dazzler, a wide host-range transposon vector for genetic transformation in vertebrates". Journal of Molecular Biology. 302 (1): 93–102. doi:ten.1006/jmbi.2000.4047. PMID 10964563.
- ^ Kurtti TJ, Mattila JT, Herron MJ, Felsheim RF, Baldridge GD, Burkhardt NY, et al. (Oct 2008). "Transgene expression and silencing in a tick cell line: A model arrangement for functional tick genomics". Insect Biochemistry and Molecular Biology. 38 (10): 963–eight. doi:ten.1016/j.ibmb.2008.07.008. PMC2581827. PMID 18722527.
- ^ Graham Lawton Why Darwin was wrong about the tree of life New Scientist Magazine issue 2692 21 January 2009 Accessed February 2009
- ^ Genomic analysis of Hyphomonas neptunium contradicts 16S rRNA gene-based phylogenetic analysis: implications for the taxonomy of the orders 'Rhodobacterales' and Caulobacteraes
- ^ Zhaxybayeva O, Gogarten JP (Apr 2004). "Cladogenesis, coalescence and the evolution of the iii domains of life". Trends in Genetics. twenty (4): 182–7. doi:10.1016/j.tig.2004.02.004. PMID 15041172.
- ^ a b c Doolittle WF (February 2000). "Uprooting the tree of life". Scientific American. 282 (2): ninety–five. Bibcode:2000SciAm.282b..90D. doi:10.1038/scientificamerican0200-90. PMID 10710791.
- ^ Woese CR (June 2004). "A new biological science for a new century". Microbiology and Molecular Biology Reviews. 68 (2): 173–86. doi:x.1128/MMBR.68.2.173-186.2004. PMC419918. PMID 15187180.
- ^ Theobald DL (May 2010). "A formal test of the theory of universal common ancestry". Nature. 465 (7295): 219–22. Bibcode:2010Natur.465..219T. doi:x.1038/nature09014. PMID 20463738. S2CID 4422345.
- ^ Huang J, Gogarten JP (2009). "Ancient factor transfer every bit a tool in phylogenetic reconstruction". Horizontal Gene Transfer. Methods in Molecular Biology. Vol. 532. Humana Press. pp. 127–39. doi:ten.1007/978-1-60327-853-9_7. ISBN9781603278522. PMID 19271182.
- ^ Davín AA, Tannier E, Williams TA, Boussau B, Daubin V, Szöllősi GJ (May 2018). "Gene transfers can engagement the tree of life". Nature Ecology & Evolution. 2 (5): 904–909. doi:ten.1038/s41559-018-0525-3. PMC5912509. PMID 29610471.
- ^ Wolfe JM, Fournier GP (May 2018). "Horizontal gene transfer constrains the timing of methanogen development". Nature Ecology & Evolution. 2 (5): 897–903. doi:10.1038/s41559-018-0513-7. hdl:1721.ane/118329. PMID 29610466. S2CID 4968981.
- ^ Oliveira PH, Touchon M, Cury J, Rocha EP (October 2017). "The chromosomal organisation of horizontal gene transfer in bacteria". Nature Communications. viii (1): 841. Bibcode:2017NatCo...viii..841O. doi:10.1038/s41467-017-00808-westward. PMC5635113. PMID 29018197.
- ^ Bryant DA, Frigaard NU (Nov 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends in Microbiology. 14 (eleven): 488–96. doi:x.1016/j.tim.2006.09.001. PMID 16997562.
- ^ Avrain L, Vernozy-Rozand C, Kempf I (2004). "Evidence for natural horizontal transfer of tetO gene between Campylobacter jejuni strains in chickens". Journal of Applied Microbiology. 97 (1): 134–40. doi:x.1111/j.1365-2672.2004.02306.x. PMID 15186450. S2CID 19184139.
- ^ Darkened Forests, Ferns Stole Cistron From an Unlikely Source — and And then From Each Other Archived 2016-03-07 at the Wayback Machine by Jennifer Frazer (May 6, 2014). Scientific American.
- ^ Li FW, Rothfels CJ, Melkonian M, Villarreal JC, Stevenson DW, Graham SW, et al. (2015). "The origin and evolution of phototropins". Frontiers in Plant Science. six: 637. doi:10.3389/fpls.2015.00637. PMC4532919. PMID 26322073.
- ^ Wybouw N, Dermauw W, Tirry 50, Stevens C, Grbić M, Feyereisen R, Van Leeuwen T (April 2014). "A gene horizontally transferred from bacteria protects arthropods from host plant cyanide poisoning". eLife. iii: e02365. doi:ten.7554/eLife.02365. PMC4011162. PMID 24843024.
- ^ Yong East (2011-02-16). "Gonorrhea has picked upwardly human DNA (and that'due south just the beginning)". National Geographic. Retrieved 2016-07-14 .
Further reading [edit]
- Gyles C, Boerlin P (March 2014). "Horizontally transferred genetic elements and their part in pathogenesis of bacterial disease". Veterinarian Pathology. 51 (2): 328–forty. doi:10.1177/0300985813511131. PMID 24318976. S2CID 206510894.
- – Papers by Dr Michael Syvanen on Horizontal Cistron Transfer
- Salzberg SL, White O, Peterson J, Eisen JA (June 2001). "Microbial genes in the man genome: lateral transfer or cistron loss?" (PDF). Science. 292 (5523): 1903–6. Bibcode:2001Sci...292.1903S. doi:x.1126/science.1061036. PMID 11358996. S2CID 17016011.
About 40 genes were found to exist exclusively shared by humans and bacteria and are candidate examples of horizontal transfer from leaner to vertebrates. Cistron loss combined with sample size effects and evolutionary rate variation provide an alternative, more biologically plausible caption
- Qi Z, Cui Y, Fang West, Ling L, Chen R (January 2004). "Autosomal similarity revealed past eukaryotic genomic comparison". Journal of Biological Physics. xxx (4): 305–12. doi:ten.1007/s10867-004-0996-0. PMC3456315. PMID 23345874.
- Woese CR (June 2002). "On the evolution of cells". Proceedings of the National Academy of Sciences of the United States of America. 99 (13): 8742–7. Bibcode:2002PNAS...99.8742W. doi:10.1073/pnas.132266999. PMC124369. PMID 12077305. This article seeks to shift the emphasis in early phylogenic accommodation from vertical to horizontal gene transfer. He uses the term "Darwinian Threshold" for the time of major transition of evolutionary mechanisms from mostly horizontal to generally vertical transfer, and the "origin of speciation".
- Snel B, Bork P, Huynen MA (Jan 1999). "Genome phylogeny based on gene content". Nature Genetics. 21 (one): 108–10. doi:ten.1038/5052. PMID 9916801. S2CID 10296406. This commodity proposes using the presence or absence of a set of genes to infer phylogenies, in order to avert misreckoning factors such equally horizontal factor transfer.
- "Webfocus in Nature with gratis review articles". Archived from the original on 2005-11-02.
- Patil Pb, Sonti RV (October 2004). "Variation suggestive of horizontal gene transfer at a lipopolysaccharide (lps) biosynthetic locus in Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen of rice". BMC Microbiology. 4 (1): 40. doi:10.1186/1471-2180-4-40. PMC524487. PMID 15473911.
- Jin One thousand, Nakhleh L, Snir Due south, Tuller T (November 2006). "Maximum likelihood of phylogenetic networks". Bioinformatics. 22 (21): 2604–11. doi:10.1093/bioinformatics/btl452. PMID 16928736.
- Jain R, Rivera MC, Lake JA (March 1999). "Horizontal gene transfer among genomes: the complexity hypothesis". Proceedings of the National Academy of Sciences of the United States of America. 96 (seven): 3801–6. Bibcode:1999PNAS...96.3801J. doi:10.1073/pnas.96.7.3801. PMC22375. PMID 10097118.
- Ochman H, Lawrence JG, Groisman EA (May 2000). "Lateral cistron transfer and the nature of bacterial innovation". Nature. 405 (6784): 299–304. Bibcode:2000Natur.405..299O. doi:ten.1038/35012500. PMID 10830951. S2CID 85739173.
- Preston R (July 12, 1999). "The Demon in the Freezer". The New Yorker. pp. 44–61.
Smallpox knows how to brand a mouse protein. How did smallpox larn that? 'The poxviruses are promiscuous at capturing genes from their hosts,' Esposito said. 'Information technology tells you that smallpox was once inside a mouse or some other small rodent.'
- Szpirer C, Elevation E, Couturier M, Mergeay Chiliad (December 1999). "Retrotransfer or gene capture: a characteristic of conjugative plasmids, with ecological and evolutionary significance". Microbiology. 145 ( Pt 12) (Pt 12): 3321–3329. doi:x.1099/00221287-145-12-3321. PMID 10627031.
- "Tin transgenes from genetically modified plants be absorbed by micro-organisms and spread in this way?". GMO Safe: Results of research into horizontal gene transfer. Archived from the original on 2011-07-21.
- Whitaker JW, McConkey GA, Westhead DR (2009). "The transferome of metabolic genes explored: analysis of the horizontal transfer of enzyme encoding genes in unicellular eukaryotes". Genome Biological science. x (4): R36. doi:10.1186/gb-2009-10-four-r36. PMC2688927. PMID 19368726.
External links [edit]
- Citizendium:Horizontal gene transfer
- Citizendium:Horizontal gene transfer in prokaryotes
- Citizendium:Horizontal cistron transfer in plants
- Citizendium:Horizontal factor transfer (History)
pittmanwillieret1950.blogspot.com
Source: https://en.wikipedia.org/wiki/Horizontal_gene_transfer
0 Response to "What is genetic material transferred to an organism?"
Post a Comment